IgG4
液性免疫の担い手となる抗体は病原体の侵入からの防御に重要ですが、病気を引き起こすこともあります。抗体は、Fab(抗原結合断片)領域を通じて病原体の特定の構造を認識して、結合します。抗体は病原体の中和を仲介するだけでなく、病原体のオプソニン化も引き起こします。オプソニン化とは、抗体のFc領域を介して、マクロファージ、好中球、ナチュラルキラー細胞などの免疫細胞と相互作用し、抗体依存性細胞媒介性細胞毒性(ADCC)または抗体依存性細胞貪食(ADCP)を引き起こし、病原体を排除する仕組みです。ヒトでは、Fc領域の違いにより五つの抗体(免疫グロブリン)があります:IgM、IgD、IgE、IgA、IgG。IgM抗体は、一次獲得免疫応答の初期段階で生成されます。IgMとIgDは、ナイーブB細胞のB細胞受容体を形成します。一次IgM応答の後には、IgGまたはIgA抗体の二次的で持続的な増加が起こり、IgAは粘膜免疫応答に関与しています。IgEは寄生虫に対する防御として進化し、さまざまなアレルギー疾患にも関与しています。
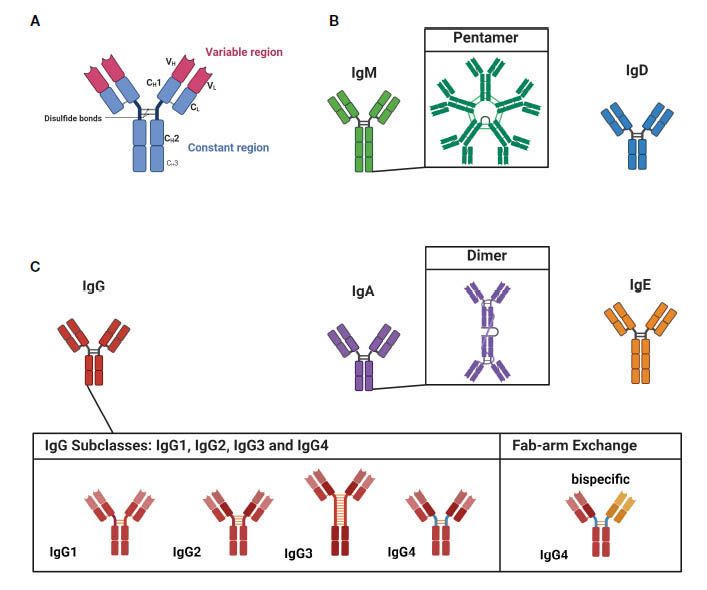
図:アイソタイプ、IgGサブクラスの構造。 文献2)より
(A) 抗体は、可変領域(VHおよびVL)として知られる2つの同一の抗原結合部位からなる二量体構造です。この領域は、Fab(Fragment-antigen Binding)領域と呼ばれ、軽鎖と重鎖の組み合わせによって形成され、抗体の抗原結合部位です。ヒンジ領域は、Fab領域を定常領域(Fc領域、CH1-CH3)に接続し、抗体のアイソタイプと機能を決定します。
(B) IgMおよびIgDのアイソタイプは、未成熟な、または「ナイーブ」な細胞と関連しています。IgMは五量体を形成します。
(C) IgG、IgA、およびIgEは、より成熟した抗体に結合しています。IgAは二量体構造を形成する能力を持っています。IgGにはIgG1、IgG2、IgG3、IgG4の4つのサブクラスがあります。IgG4サブクラス抗体は、Fab-arm交換を受けることができ、二重特異性を持つことができます。
IgG抗体には4つのサブクラスがあり、血液中の濃度が高い順に番号が付けられています。血液中の濃度が最も高いサブクラスはIgG1(血液中の濃度:60%)で、オプソニン化、ADCC、および補体活性化の作用が強く、対照的に、IgG2(血液中の濃度:32%)抗体は補体の活性化にはあまり効果がないが、カプセル化された細菌のオプソニン化には強力です。3番目に多いサブクラスはIgG3(血液中の濃度:4%)で、補体カスケードを活性化し、ADCCを誘導し、毒素を中和する作用が強く認められます。血液中の濃度が最も低いサブクラスであるIgG4(血液中の濃度:4%)について、その特徴、生理学的役割、病気との関わりについて詳しく掘り下げていきます。
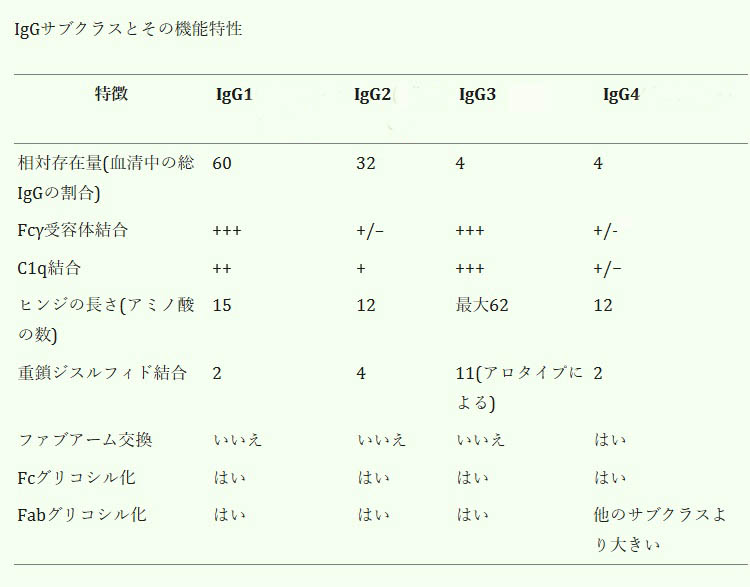
文献1)より
IgG4の特徴
IgG4は他のIgGサブクラスと比較して独自の特性を持ち、アレルギー疾患において有益な機能を持つ抗炎症性の「良性」抗体として広く認識されています。IgG4 は、他のIgG サブクラスよりもFc 受容体や補体などの多くのエフェクター分子に対する親和性が低いですが、Fab-Arm交換という独特な能力を持っています。IgGサブクラス抗体は、通常、2つの同一の結合部位を持つ二量体です。Fab-Arm交換により、IgG4サブクラス抗体は(半)分子を他のIgG4抗体と交換することができ、2つの異なる可変領域を持つ二重特異性抗体が得られます。この反応は追加のタンパク質や補因子を必要としないため、IgG4抗体がin vivoで絶えずアームを交換する非常にダイナミックなプロセスをもたらし、それがヘテロ二価IgG4抗体の絶えず変化するレパートリーを生成します。Fab-arm交換型二重特異性IgG4抗体は抗原を架橋できず、免疫複合体を形成しません。したがって、IgG4抗体は一般に免疫調節性と考えられており、「ブロッキング抗体」と呼ばれています。特に、アレルギー反応と寄生虫感染の両方において、IgG4 反応は免疫寛容を誘導し、炎症を制限します。
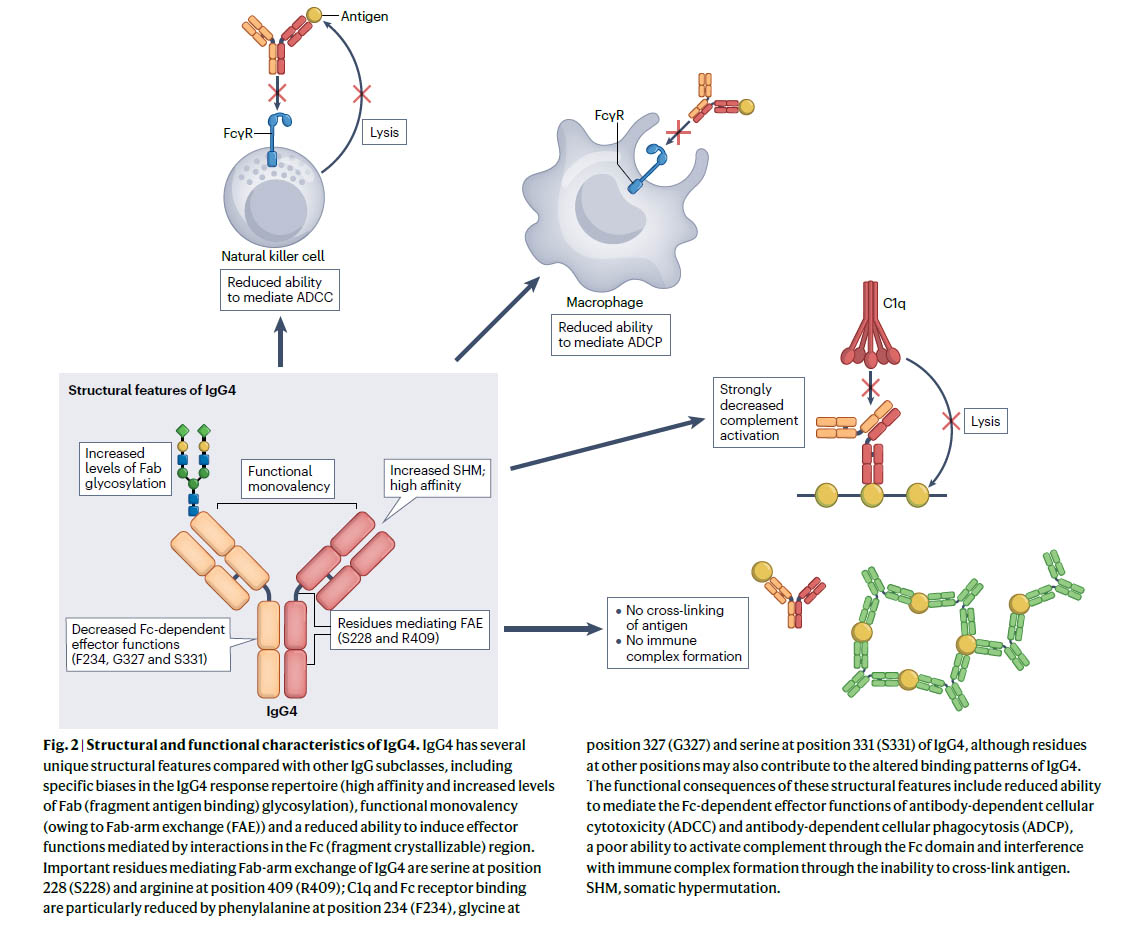
図:IgG4の構造的および機能的特性 文献1)より
IgG4は、他のIgGサブクラスと比較して、Fab-arm exchangeによる機能的一価性、Fc領域での相互作用によって媒介されるエフェクター機能の低下など、いくつかのユニークな構造的特徴を持っています。これらの構造的特徴の機能的影響には、抗体依存性細胞毒性(ADCC)および抗体依存性細胞食作用(ADCP)のFc依存性エフェクター機能を媒介する能力の低下、Fcドメインを介した補体活性化能力の低下、抗原を架橋できないことによる免疫複合体形成の妨害が含まれます。SHM、体細胞超突然変異。
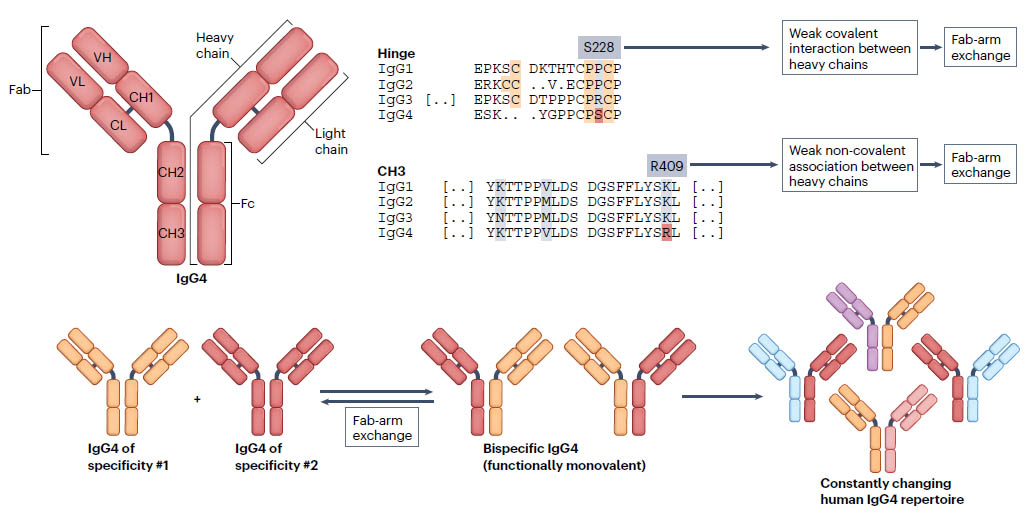
図:Fab-arm exchange 文献1)より
IgG抗体は、IgG4を除いて、通常、対称的な分子です。IgG抗体は、2つの半分子の共有二量体であり、各半分は軽鎖と重鎖で構成されています。形質細胞によって産生される抗体は、二価および一重特異性(同じ特異性の2つの抗原結合部位を有する)です。しかし、ヒトIgG4は非常に動的な抗体であり、これらの半分子は、Fab(fragment antigen binding)-arm exchangeと呼ばれるプロセスで他のIgG4半分子と解離して再結合することができます。得られたIgG4抗体は、2つの異なる抗原結合部位を持つ非対称の二重特異性構造です。血液中のIgG4の最大99%が二重特異性であると考えられています。
IgG4の生理学的役割
アレルギー
アレルギーは、発疹、腫れ、かゆみ、上気道過敏症、重症の場合はショックなどの症状を引き起こす過敏反応を特徴とします。これらの反応は、TH2細胞によるIL-13およびIL-4の放出と、それに続くB細胞のIgE産生へのクラススイッチを引き起こすアレルゲンによって生じます。アレルゲンに結合したIgEによるFcεRI刺激に反応して、肥満細胞と好塩基球はヒスタミン、サイトカイン、ケモカインを放出し、血管系と組織に影響を及ぼして過敏症状を引き起こします。アレルゲン特異的IgG4反応は、草やシラカバの花粉、猫、ハチ毒、ピーナッツ、牛乳など、さまざまなアレルギーで報告されています。アレルゲン特異的IgG4の力価の上昇は、一般的にアレルギーに対する耐性の増大および過敏症症状の軽減と相関しています。IgG4を介したアレルギー症状の予防は、少なくとも3つの作用機序の結果であると考えられています。アレルゲン結合の競合によるIgEの活性阻害および肥満細胞と好塩基球の脱顆粒の防止、B細胞および樹状細胞上のIgEによるT細胞への抗原提示の阻害、IgG4の機能的一価性による免疫複合体の形成の防止です。アレルゲン特異的IgG4反応の誘導は、アレルゲンへの長期曝露およびIL-10産生の増加に起因すると考えられています。IL-10はIgE分泌を減少させ、IgG4産生を増加させることが示されております。
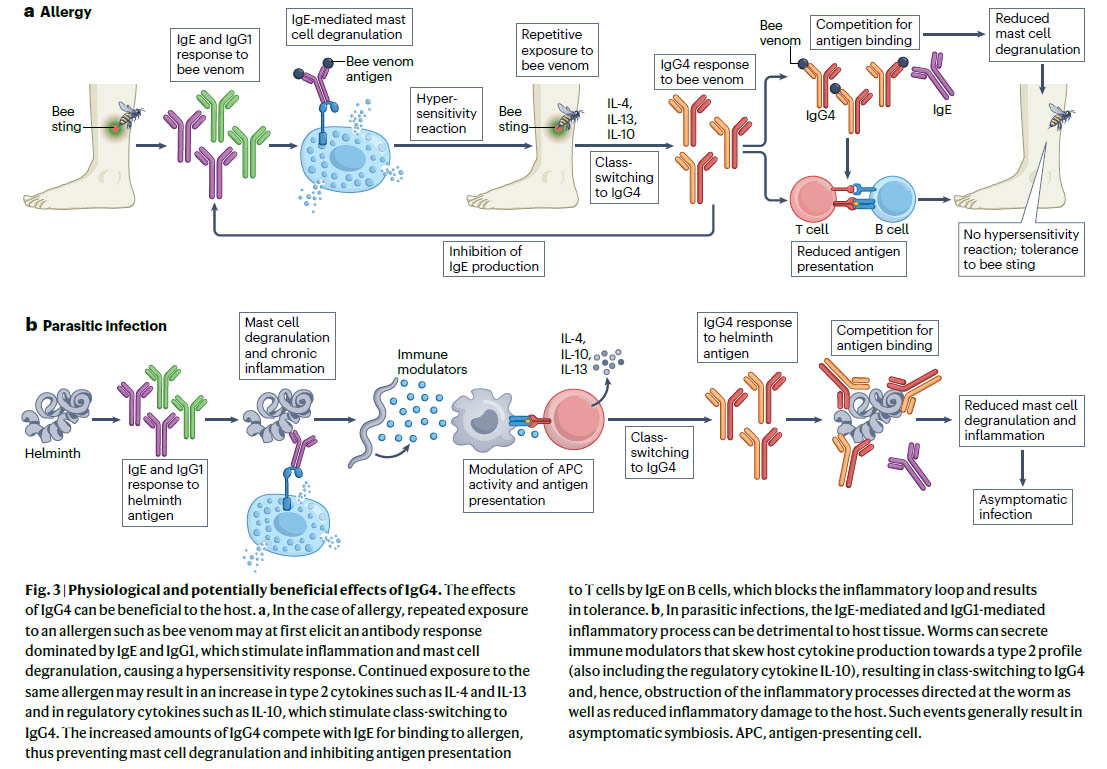
図:IgG4の生理学的および潜在的に有益な効果 文献1)より
a. アレルギーの場合、ハチ毒などのアレルゲンに繰り返し曝露すると、最初はIgEやIgG1が優勢な抗体反応を誘発し、炎症や肥満細胞の脱顆粒を刺激し、過敏反応を引き起こす可能性があります。同じアレルゲンへの継続的な曝露は、IL-4やIL-13などの2型サイトカインやIL-10などの調節性サイトカインの増加をもたらし、IgG4へのクラス切り替えを刺激する可能性があります。IgG4の量の増加は、アレルゲンへの結合をめぐってIgEと競合するため、肥満細胞の脱顆粒を防ぎ、B細胞上のIgEによるT細胞への抗原提示を阻害し、炎症ループをブロックして耐性をもたらします。
b、寄生虫感染症では、IgE媒介性およびIgG1媒介性の炎症過程が宿主組織に有害である可能性があります。線虫は、宿主サイトカインの産生を2型プロファイル(調節性サイトカインIL-10も含む)に偏らせる免疫調節物質を分泌し、その結果、IgG4にクラスが切り替わり、線虫に向けられた炎症過程が妨げられ、宿主に対する炎症性損傷が減少します。このようなイベントは、通常、無症候性の共生をもたらします。APC、抗原提示細胞。
寄生虫感染
IgG4反応は寄生虫感染時にも発生する可能性があります。宿主は通常、寄生虫に対してB細胞介在性およびT細胞介在性の広範な免疫反応を発現します。宿主の免疫反応を回避するために、寄生虫はIL-10などのサイトカインの産生と Treg 細胞の誘導を刺激します。その結果、一部の患者では、抗寄生虫 B 細胞反応が IgG4 へのクラススイッチ(IgG反応の最大90%がIgG4)を起こし、症状を起こさないと考えられています。実際、フィラリア抗原で攻撃されたフィラリア症患者のIgEオプソニン化好塩基球によるヒスタミン放出は患者由来のIgG4によって阻害され、ヒスタミン放出はIgG4レベルと逆相関していました。興味深いことに、寄生虫によるIgG4媒介宿主免疫の減弱の結果、自己免疫疾患やアレルギーから宿主が保護される可能性があることが示唆されています。このような役割は、「衛生仮説」と一致します。この仮説では、高所得国では寄生虫への曝露が少なく、衛生基準が向上しているため、自己免疫疾患やアレルギー疾患の有病率が上昇する可能性があると提唱されています。
IgG4の関連する病態・疾患
IgG4の特異的欠損は、単独の現象として(症例の約30%)、またはIgG2、IgA、IgG1など他の抗体サブクラスの欠損と組み合わせて発生する可能性があります。IgG4の選択的欠損またはIgG4レベルの大幅な低下は極めてまれです。一部の個人(主に小児)では、IgG4欠損は、再発性呼吸器感染症、アレルギー、カンジダ症、慢性下痢、慢性真菌感染症に関連しています。また、最近の研究では、炎症性腸疾患患者の約20% にIgG4 欠損が見られ、炎症性腸疾患の増悪と関連していることが示されています。IgG4 高ガンマグロブリン血症は、健康な人口の約5% に発生しますが、無症状のことが多いと報告されています。その他下記に示す病態、疾患が、病原性IgG4反応に関連しています。
抗腫瘍反応
抗腫瘍抗体は、腫瘍細胞に結合し、抗体依存性細胞媒介性細胞毒性(ADCC)、抗体依存性補体沈着、抗体依存性細胞貪食(ADCP)を刺激することで、悪性腫瘍を抑制または排除することができます。しかし、1977年に黒色腫患者を対象とした前向き研究で、IgG4の高レベルが生存に悪影響を及ぼすことが確認されました。それ以来、一部の悪性腫瘍はIgG4に対する抗腫瘍抗体反応のクラススイッチを誘導することで宿主の免疫防御を回避することが明らかになっています。IgG4は、腫瘍抗原への結合をめぐって他の抗体(サブ)クラスと競合し、その抗炎症特性により抗腫瘍免疫反応の誘導を阻害します。免疫反応がない場合、腫瘍細胞の増殖および転移能力が高まり、病気の進行と生存率の低下につながります。 IgG4へのクラススイッチによる免疫回避は、黒色腫、胆管癌、結腸癌、膵臓癌、神経膠芽腫の患者で観察されています。悪性腫瘍患者の血清では、IgG4の総レベルとIgG4+ B細胞数の両方が増加する可能性があり、これは予後不良の指標です。
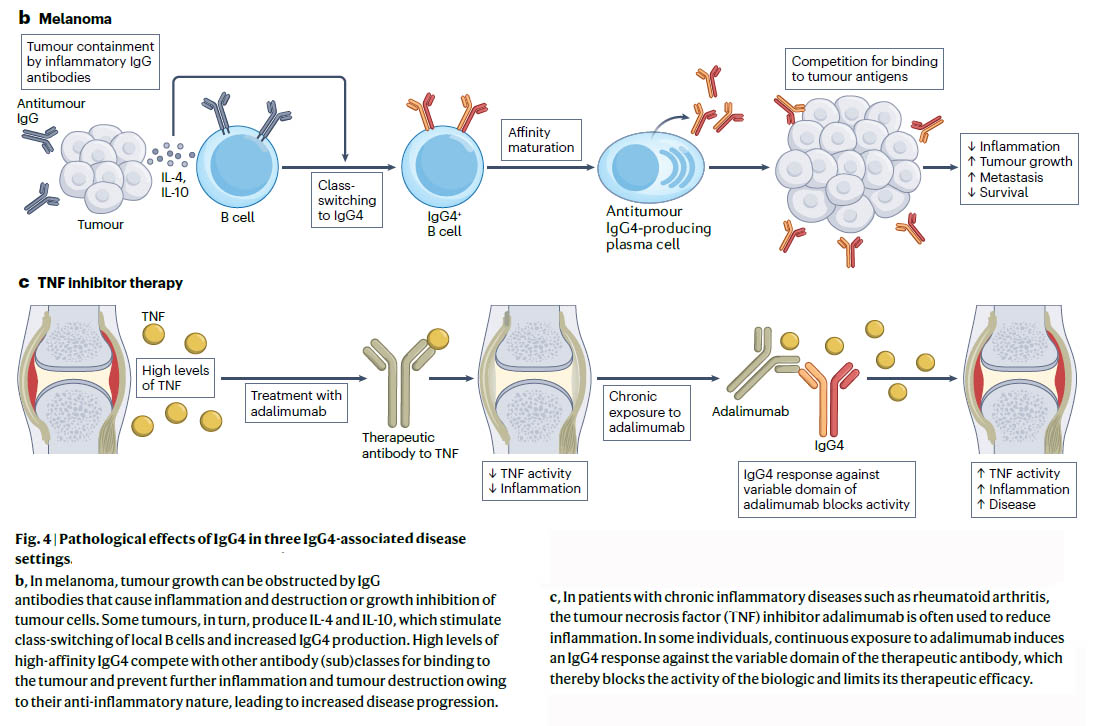
図:IgG4関連疾患におけるIgG4の病理学的影響 文献1)より
b.黒色腫では、腫瘍細胞の炎症や破壊、または成長阻害を引き起こすIgG抗体によって腫瘍の成長が妨げられることがあります。一部の腫瘍では、IL-4とIL-10が産生され、これが局所B細胞のクラス切り替えを刺激し、IgG4産生を増加させます。高レベルの高親和性IgG4は、腫瘍への結合をめぐって他の抗体(サブ)クラスと競合し、その抗炎症性によりさらなる炎症や腫瘍の破壊を防ぎ、疾患の進行を増加させます。
c. 関節リウマチなどの慢性炎症性疾患の患者では、腫瘍壊死因子(TNF)阻害剤であるアダリムマブが炎症を軽減するためによく使用されます。一部の個人では、アダリムマブへの継続的な曝露により、治療用抗体の可変ドメインに対するIgG4応答が誘導され、それによって生物学的製剤の活性が阻害され、その治療効果が制限されます。
抗生物学的反応
IgG4偏向反応は、生物学的療法への慢性的な曝露の結果として発生することもあります。このような反応は、先天性血友病Aまたは血友病Bの治療に使用される凝固因子FVIIIおよびFIX、多発性硬化症の治療に使用されるインターフェロンβ、関節リウマチやクローン病などの炎症性疾患の治療に使用される腫瘍壊死因子(TNF)阻害剤アダリムマブおよびインフリキシマブについて報告されています。これらの抗生物学的反応の結果、生物学的製剤の治療効果が損なわれてしまいます。すべての生物学的製剤がIgG4偏向抗薬物反応を引き起こすわけではありません。特定の生物学的製剤がこれらの反応を引き起こし、他の生物学的製剤が引き起こさない理由は不明です。
IgG4自己免疫疾患(IgG4-AID)
IgG4関連疾患
文献
1)The unique properties of IgG4 and its roles in health and disease. nature reviews immunology 2022 https://doi.org/10.1038/s41577-023-00871-z
2)Exploring the depths of IgG4:insights into autoimmunity and novel treatments. Front. Immunol. 2024, 15:1346671. doi: 10.3389/fimmu.2024.1346671
<2025年3月22日作成>
IgG4記事一覧
IgG4自己免疫疾患
IgG4-AIDは、2015年に抗体が媒介する自己免疫疾患の別のサブグループとして初めて定義されました。IgG4-AIDは、既知の抗原に対する主にIgG4サブクラスの自己抗体反応を特徴とします。これらの疾患は、標的抗原の主な作用部位に応じて、腎臓、中枢神経系/末梢神経系、造血系、皮膚など、多くの臓器...